A Fundamental Unit Of The Brain And Its Functional Mechanisms
Background
Neuron
A neuron is the functional unit of the brain. The human brain contains almost 100 billion neurons (2). The most simplified version of a neuron is composed of a cell body and several projections toward other neurons. A neuron receives and sends information in the form of action potential through these connections. It includes two classes of neurons: excitatory and inhibitory. The excitatory neurons send positive inputs to other neurons, which depolarise their membrane potential and prime the activation. The inhibitory neurons send negative inputs, which hyperpolarise the membrane potential and reduce the likelihood of activation. If a neuron is stimulated enough, proving its membrane potential to exceed the membrane threshold the neurons fires. If its membrane potential does not reach that threshold, the neuron stays inactivated. Hence, the summation of excitatory and inhibitory inputs provides a neuron to fire or not.

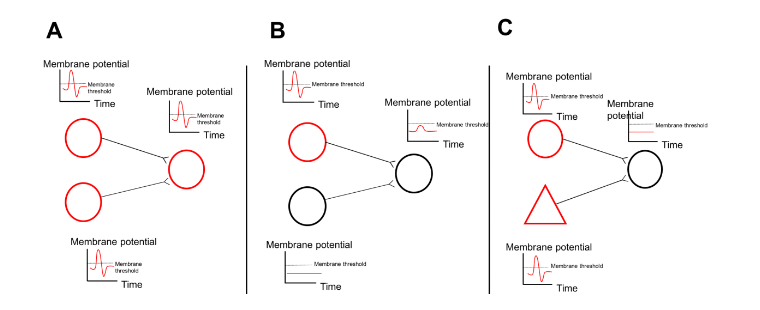
Figure 2: Activation of a neuron. A circle represents an excitatory neurons, a triangle represents an inhibitory neurons. A. The two excitatory neurons on the left fire send an action potential to the third one on the right. Its membrane potential reaches the membrane threshold leading to its activation. B. In this case, only one excitatory neuron fires the membrane, potential of the neuron on the left increases but it does not reaches the membrane threshold, it therefore stays inactivated. C. The excitatory and inhibitory neurons fire provides an excitatory and an inhibitory input. The summation of inputs is therefore zero and the membrane potential of the third neuron remains unchanged.
Hebbian learning
A neuron makes an average of 103 connections, which can vary in strength. The strength of connections between two neurons represents the amount of influence one neuron. In 1949, Donald Hebb proposed that, two neurons, which are activated at the same time, increase the connection between them (3). In a network with many connections, the Hebbian rules provide the strengthening of all the connections, leading to a hyper connected network. Under these conditions, the stimulation of a single element would activate the entire network. A new theory allows more flexibility for the network. In this revision, two neurons still increase their connections when they fire at the same time. However, the connection decreases, when one fire and the other does not. Hence, the number and the strength of connection for the neurons can be increased and decreased. The synchronisation of firing is therefore the key for the evolution of connections. This theory has then evolved into the spike-time-dependency (4), which has been observed in the Optic Tectum of Xenopus Laevis (5).

Figure 3: A. Hebbian Learning. The repeated synchronisation of firing of neurons provides the strength of connection. B. Spike-time-dependency. The repeated de-synchronisation of firing of neurons decreases the strength of connection
Cell assembly
The repeated co-activation of neurons provides the strengthening of their synapses, leading to the emergence of a distributed network, called cell assembly. Since, neurons of a cell assembly are strongly connected with the stimulation of a fraction of them that leads to the activation of the whole population or at least a consequential part of it. This process is called ignition (6). After an ignition, neurons, which are part of the cell assembly, start their refractory period. Neurons were not activated at the same time, therefore, their deactivation is not synchronised too. Since, they are strongly connected and some of them can be reactivated by other that fired before. Thus, the activity is retained and the deactivation of the cell assembly decreases slowly. This state is called reverberation. It has been observed in monkeys (7) and rats (8) the activity can last for ten seconds in absence of stimulus. The observation of an object triggers the activation of neurons in the visual area. When, it occurs several times, these neurons reinforce their connections. The cell assembly of neurons may therefore be considered as the cortical representation of an object. However, all neurons of a cell assembly do not have to be active to provide its mental representation. It is assumed that, a certain proportion of active neurons in the cell assembly are sufficient. This proportion is called ignition threshold and it is considered as the point of no return, after which the ignition cannot be stopped.

Figure 4: A. Emergence of the cell assembly. The repeating co-activation of neurons leads to the emergence of a cell assembly B. Ignition of a cell assembly. The stimulation of one neuron of the cell assembly provides the ignition of the cell assembly. C. The reverberation of a cell assembly after its ignition.
The strength of connection is not uniform among the neurons in a cell assembly. Some neurons exhibit a high correlation among each other, whereas others have weaker connections. It therefore is possible to differentiate the core, in which the connections are the strongest and the periphery in which the connections are weaker. Nevertheless, the distinction between the core and the periphery is arbitrary.

Word as a cell assembly
An object such as a phone is multi-modal. In addition to its visual features it includes auditory ones. Neurons involved in the cell assembly for phone are therefore distributed across the auditory and the visual cortex. Spoken language involves auditory and motor areas. Thus, the word phone has a spread cell assembly through several regions of the brain. These multimodal activations have been observed in imagery studies (9) (10) (11). The theory of action-perception circuit as a basis of language has then been elaborated (12).
Furthermore, some words have the exact same pronunciation such as knight and night. In case of homophones, one input triggers the activation of the same neurons in the two cell assemblies, but only one activation is possible there. Hence, there is an overlapping of cell assemblies in the auditory area. There is therefore a race between the two cell assemblies, which ends in the ignition of one of them, the first to reach the ignition threshold. The outcome of the race depends on the number of inputs received as well as the strength and number of connections in the cell assembly.
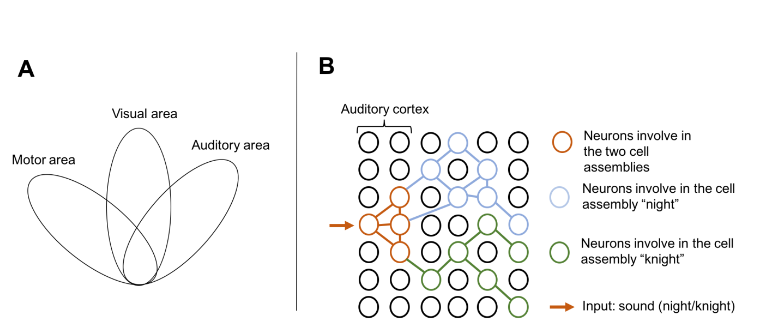
Figure 6: A. Example of a distributed cell assembly across visual motor and auditory areas. B. The race between two cell assemblies in case of homophones. In this example, the cell assembly for night receives two inputs and has more connections than the cell assembly for knight. Hence, night should be the first to reach the ignition threshold.
Sentence as a chain of cell assemblies
Syntax
In linguistic “syntax” is the set of rules, which governs the words in a sentence. Words are classified into lexical categories. These classes correspond to traditional part of speech such as noun, verb or determinant. Syntax also includes word order. For instance, “the cat eats the mouse” is a correctly syntactic sentence, “whereas” cat the eats mouse the” is not. A noun can be preceded by a determinant, but the opposite is not true. Syntax is therefore a major component of grammar and the relations between the words. In the same example, “the cat eat the dog” is not syntactically correct. The third person requires the suffix “s” in English. It is therefore necessary to maintain information from the first word to ensure the correct relation with the second. This is called grammar dependency. Moreover, these rules hold, when two words are spaced by others. For example, in the sentence “the cat that you bought me for my last birthday eats the mouse”, 8 words intervenes between the interdependent words “cat” and “eats”. This very distant relationship between two elements is called long term dependency.
Evidences from songbirds
In a sequence of two words, A and B the cell assembly α representing the word A is initially activated, followed by the cell assembly β representing the word A. Recording such cell assemblies in human is
currently impossible for ethical and technical reasons. However, recent progress in large scale recording has provided the opportunity of studying cell assemblies in animal. Vocalisation of songbirds is strongly related to human language. It is also composed of sequences of syllables and their acquisition is similar to human infants (13). In addition, songbirds would process syntax. They are able to exclude grammatical patterns and classify grammatical ones (14). Sequential activity of neurons has been observed in high vocal centre of Zebra finch (15). It has been proposed that, vocalisation is the result of the activation of two chains of neurons each representing a syllable (16) (17). The first chain of neurons primes the activation of the second. This mechanism is called synfire chain and it could be the basis of serial order in the brain such as syntax (18).
Synftre chains
A synfire chain makes a bridge between two cell assemblies. The cell assembly α activates the synfire chain which would prime the activation of the second cell assembly β (19). The size of a cell assembly is still not yet knows. However, studies in animal showed that, an average of 150 to 300 neurons oscillated in the same cycle and it could be part of the same cell assembly (20) (15). In order to prime the activation of a cell assembly of this size, a synfire chain cannot be composed of a little number of neurons. Hence, a synfire chain consists of consequent sets of neurons defining a spatio-temporal pattern. Thus, a synfire chain could explain how a noun is primed by the activation of determinant. Nevertheless, a synfire chain is bi-directional. It can be activated by the cell assembly α, but also by the cell assembly β. Since a noun cannot be followed by a determinant, this non-specificity is less likely to explain syntax. A different mechanism might be involved.
Sequence detectors
Friedmann Pulvermller proposed that, a third neuronal element Ω is involved in the sequence A-B. This third entity is called sequence detectors and it would mediate the transition from one cell assembly α to another β, but not in the opposite direction (21). Such sequences would have been found for movement detection in the visual system of insects (22) 1 but also in the retina of mammals involving inhibitory
loops (23). These sequence detectors may be considered as the neuronal grammar in the brain. Indeed, if a sequence Ω mediates the transition from α1 to β1, it is also possible that, it mediates the transition from α2 to β2. In this case, α could represent the lexical class determinant and β the lexical class noun. Thus, the repetition in sequence of word pairs strengthens the sequence detectors, which would represent a specific syntactic rule. A new input or word learnt, would therefore link to this sequence detectors, establishing its syntactic relationship.
The sequences of determinant-noun or noun-verb might create an overlap of words of the same class. This would constitute a lexical cell assembly, which is directly connected to the sequence detectors. Studies in imagery have not shown such a lexical overlap. This might be explained by the impossibility of distinguishing the semantic from lexical part of a word.
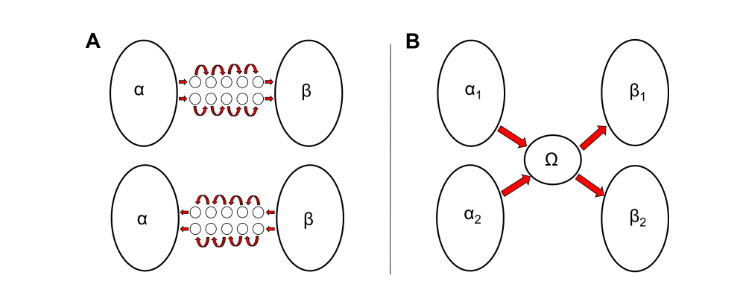
Figure 7: A. Bi-directionality of synfire chains. On the top, it is the sequence A-B. The activation of the cell assembly α activates the synfire chain which primes the activation of the cell assembly β. Below, the sequence B-A. The activation of the cell assembly β activates the synfire chain which primes the activation of the cell assembly α. B. Sequence detectors. The cell assembly α1 or α2 activates the sequence detectors Ω which primes the activation of the cell assembly β1 or β2. α and β represent two distinct lexical categories.
The mechanism of these sequence detectors remains unclear. They would be an ensemble of neurons which performs the specificity of the sequence by unequal weights between the two entities they mediate. Thus, if the sequence detector Ω is specific of the sequence A-B, it is less connected to α than it is to β. In this way, only the activation of α following by the activation of β fully activates Ω. Whereas, the sequence B-A which α is preceded by β fails to activate Ω. This is due to the non-linearity decline of activities, which makes the summation of activation unequal in one sequence to the other.
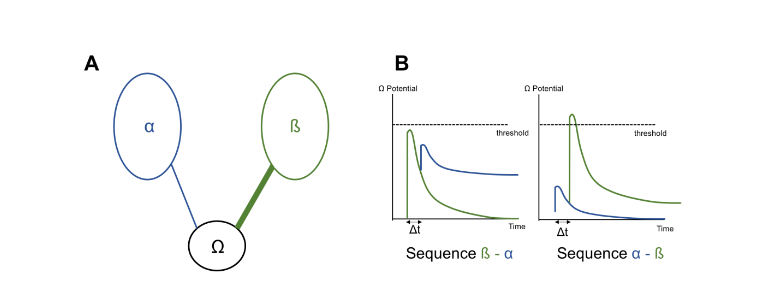
Figure 8: A. The sequence detectors Ω, with weaker weight for α than β
B. The sequence β then α does not trigger the activation of the sequence detectors Ω. Whereas the sequence α then β does.
These asymmetrical weights would be based on the different distances between α, Ω and β. Neurons in the same column are closer and therefore it is more likely to present strong connections, when they are active at the same time. While, the connections from one column to another are weaker (24). Thus, the cell assembly β which is strongly connected to the sequence detector would be in its vicinity and more particularly in the halo of β (21).

Figure 9: Representation of sequence detectors. The cell assembly α1 is weakly connected to the sequence detector representing by α1β1 in the halo of the cell assembly β1. The sequence detector α1β1 is strongly connected to the cell assembly β1. The cell assembly β1 is also connected to the sequence detector β1α1 but the non occurrence of the sequence B1A1 would prune such connection.
Moreover, these connections are also known as reciprocal (25) (26), so how α1 could specifically prime β1?
For the sequence A1B1:
First, A1 occurs which triggers the ignition of the cell assembly α1. Because α1β1 was totally inactive and is weakly connected to α1 it stays inactive.

α1 starts to be deactivated, it enters in its reverberating phase. Therefore, it continues to send activation to the sequence detector α1β1 which is strongly connected to β1 and are therefore now both primed. Note that β1 is also primed by the connection from α1 which are in its halo
Then, B1 occurs leading to the ignition of β1 which will fully activate the sequence detector α1β1. Since, the sequence detector is weakly connected to α1 which has already ignited, it is not primed.
In the opposite sequence B1A1:
B1 occurs and triggers the ignition of β1 which consequently primes the sequence detector α1β1.
β1 is deactivated and enters in its reverberating phase. Therefore, it continues to send activation to the sequence detector α1β1. However, α1 is not primed because the connections from β1 are weak.
A1 occurs which triggers the ignition of the cell assembly α1 but not of the sequence detector α1β1.
2.4 Aim of this study
In order to prove the existence of the sequence detectors it would necessitate a recording of thousands of neurons, including the estimation of their weights. The current techniques are far from this expectation (27), hence other means are crucial. Artificial neural network based on neuro-anatomical and neuro- functional principles are a new resource for these kinds of experiments. Thus, sequence detectors have already been analysed in a computational model, in which unequal weights were implemented (28). The authors found that, cell assemblies for an abstract lexical class were specifically primed if another class were previously activated. Although, this supports the theory of sequence detectors, the network was pre-structured to demonstrate the veracity of this mechanism. The goal of this study was therefore to explore the formation of sequence detectors using a computational model which was not pre-determined to observe them. This model is based on Hebbian learning and has already shown the formation of cell assemblies in language (1). Thus, this model has been trained with abstract word in sequence. The goal was to observe and understand the possible formation of sequence detectors.
Continue your exploration of A Comprehensive Overview of its Scope and Role with our related content.



- 24/7 Customer Support
- 100% Customer Satisfaction
- No Privacy Violation
- Quick Services
- Subject Experts